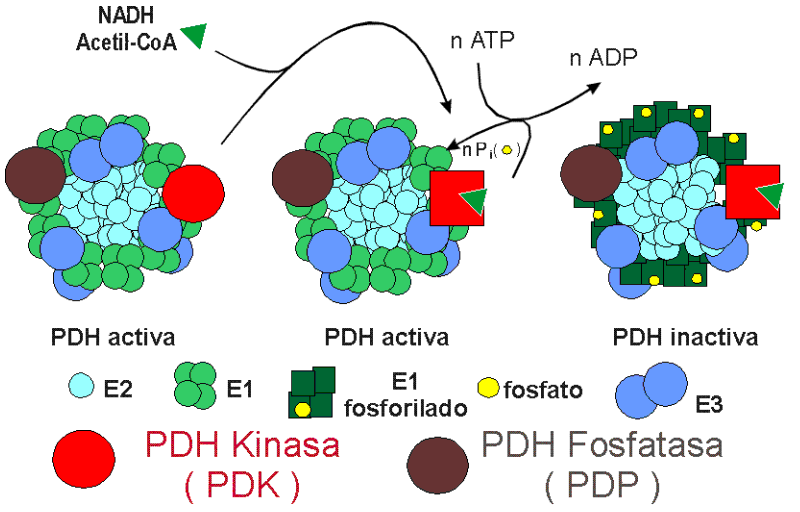
INACTIVACIÓN DEL COMPLEJO PDH
El complejo Piruvato-Deshidrogenasa ( PDH ) presenta una regulación estricta por Acetil-CoA y NADH que inhiben competitivamente el sistema multienzimático activo.
Recordemos que Acetil-CoA y NADH son productos de la reacción catalizada por PDH :
Piruvato + CoA + NAD+ => Acetil-CoA + NADH + CO2
El complejo enzimático Piruvato-Deshidrogenasa ( PDH ) puede encontrarse activo e inactivo. La inactivación es llevada a cabo por una Proteina-Kinasa ( PDH-Kinasa o PDK ) Mg+2-ATP dependiente unida al complejo PDH. Esta Kinasa actúa fosforilando tres restos de Serina de las subunidades "alfa" del Enzima 1 ( E1 ) del complejo PDH.
La PDH-Kinasa también se encuentra fuertemente regulada. La regulación del complejo PDH depende de la regulación de :
a) La PDK ( PDH-Kinasa ), inductora de la fosforilación del complejo Piruvato-Deshidrogenasa y por tanto de la inactivación del complejo PDH.
b) La Proteína-Fosfatasa ( PDH-Fosfatasa o PDP ) inductora de la defosforilación de la Piruvato-deshidrogenasa y por tanto de la activación del complejo PDH.
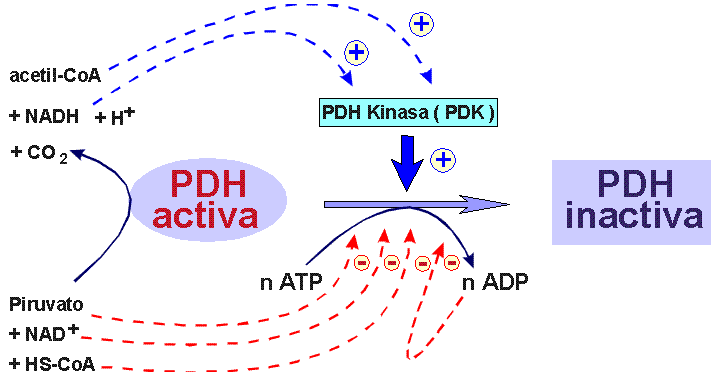
La PDH-Kinasa ( PDK ) se activa por los productos de la reacción catalizada por el complejo multienzimático Piruvato-deshidrogenasa, que son el Acetil-CoA y el NADH. estos productos al activar a la PDH-Kinasa inducen fosforilación del complejo PDH y por tanto su inactivación.
Además, si analizamos el control de las deshidrogenasas que participan en el ciclo de Krebs ( CTC ) y producen NADH ( isocitrato DH, a-cetoglutarato DH y malato DH ), podemos constatar que al igual que la PDH estas tres Deshidrogenasas son inhibidas por su producto NADH :
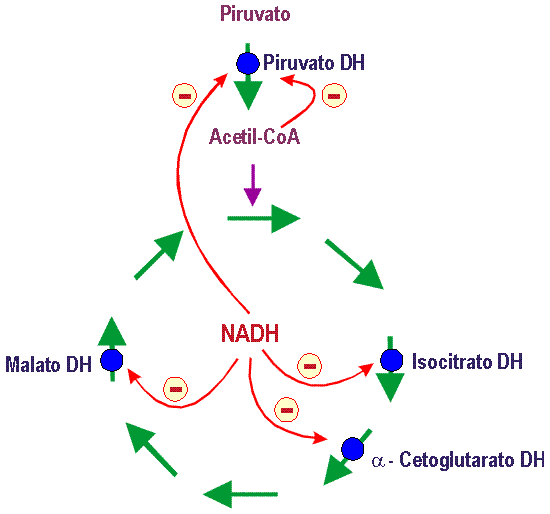
Por tanto el NADH inhibe las cuatro Deshidrogenasas productoras de NADH ( inhibición por el sustrato ) : Piruvato DH, isocitrato DH, a-cetoglutarato DH y malato DH. Esta inhibición en el caso de la PDH se realiza a través de la proteína reguladora PDH-Kinasa ( PDK ), que modula la inactivación de la PDH.
Tanto el NADH como el Acetil-CoA ven incrementados sus niveles en mitocondria durante la b-oxidación de los ácidos grasos, con paralela disminución en los niveles mitocondriales de NAD+ y de CoA-SH. La b-oxidación de los ácidos grasos es la principal fuente de Acetil-CoA en la mitocondria, que mediante su incorporación al CTC conduce a elevados niveles de NADH y FADH2 que se integraran en la cadena transportadora de electrones y fosforilación oxidativa para conducir finalmente a elevar los niveles de ATP, con gasto de O2.
En hígado, niveles elevados de cAMP pueden tambien ser empleados en la síntesis de cuerpos cetónicos tales como acetoacetato y b-hidroxibutirato.
La PDH-Kinasa ( PDK ) se inactiva por CoA-SH, ADP, NAD+ y Piruvato. Cuando se produce una elevación en los niveles tisulares de piruvato, por ejemplo en citosol por activación de la glicolisis, el piruvato entra en el interior de la mitocondria mediado por su transportador específico, y se inhibe la PDH-Kinasa ( PDK ) y por tanto el complejo PDH ( presente solo en mitocondria ) alcanza su máxima actividad.
La PDH en mitocondria de tejido adiposo es también activada por Insulina, mientras que en tejido cardiaco la Adrenalina ( Epinefrina ) activa el complejo Piruvato-DH. Los mecanismos por los que estas hormonas realizan su función reguladora sobre la actividad enzimática son desconocidos. Solo se conoce que tales efectos no pueden correlacionarse con alteraciones en los niveles de cAMP, ya que tanto la PDH-Kinasa ( PDK ) como la PDH-Fosfatasa ( PDP ) reguladoras de la actividad PDH, son insensibles a cAMP.